

 Наука в практике аквариумного рыбоводства
Наука в практике аквариумного рыбоводства
Подавляющее большинство разновидностей аквариумных рыб выведено не специалистами, а аквариумистами-любителями. Практически каждый аквариумист, основательно освоивший разведение того или иного вида и приступивший к массовому разведению, вольно или невольно становится селекционером, особенно, если полюбившиеся ему рыбки остаются в его домашнем водоеме не один год и не одно поколение. Получив многочисленное потомство, аквариумист оставляет у себя на племя определенное количество самых лучших с его точки зрения производителей с понравившимися ему формой плавников и окраской. Если такая отбраковка осуществляется в течение ряда поколений в одном и том же направлении, то интересующие аквариумистов признаки не только сохраняются, но и улучшаются. Такой метод сохранения и улучшения пород называют массовым отбором.
Массовый отбор может проводиться бессознательно, если селекционер не преследует достижения заранее поставленной цели, или сознательно, если он стремится к определенному эталону: выводит, например, вуалехвоста с более длинным, тонким и прозрачным хвостовым плавником.
Заметив среди выращенных рыбок хотя бы одну, существенно отличающуюся от своих братьев и сестер более красивой формой плавников или окраской, аквариумист оставляет ее на племя и старается спарить с особью противоположного пола—носительницей признака. Такой метод селекционеры называют индивидуальным отбором.
Индивидуальный отбор дает лучшие результаты по сравнению с массовым, иногда чередуется с ним. Однако осуществление индивидуального отбора не всегда возможно, так как редко удается подобрать пару производителей — носителей нужного признака. Чтобы закрепить его, приходится получать потомство от производителей, из которых только один (самец или самка) обладает желательным признаком.
Получив потомство от непохожих друг на друга родителей, в первом поколении в подавляющем большинстве случаев не удается обнаружить ни одного потомка, похожего на своего отца (мать) — носителя нужного признака.
Тогда обладателя нового признака (если это самец) приходится спаривать с дочерями или с сыновьями (если обладатель нового признака — самка).
Носителя одного или нескольких привлекательных для селекционера признаков называют выдающимся производителем. Чем больше поколений удается получить от выдающегося производителя, отбирая в каждом поколении потомков наиболее похожих на своего предка, тем больше шансов закрепить полюбившийся признак. Такой метод селекционеры называют инбридингом на выдающегося производителя. Если выдающийся производитель погиб, приходится получать потомство, спаривая его потомков — братьев с сестрами.
В обоих случаях мы вынуждены прибегать к нежелательному близкородственному разведению — инбридингу, с последствиями которого — инбредной депрессией (вырождением, снижением жизнестойкости, плодовитости, темпа роста и т. п.) — придется бороться, применяя специальные приемы разведения.
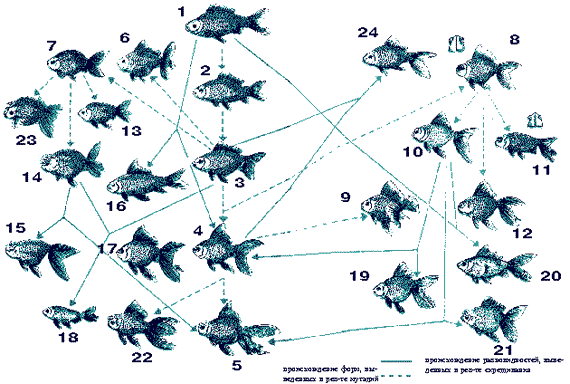
Генеалогия основных разновидностей золотой рыбки, созданных китайскими и японскими рыбоводами ( по Матсуи): 1 - фуна, исходная форма, дикий карась; 2 - хибуна, обыкновенная золотая рыбка, ярко-красная; 3 - вакин, японская золотая рыбка; 4 - риукин, вуалехвост; 5 - оранда,львиная голова (голландская); 6 - юкин, павлиний хвост; 7 - маруко, без спинного плавника; 8 - красный телескоп; 9 - тоса; 10 - калико пестрый, пестрый телескоп; 11 - небесное око; 12 - черный телескоп; 13 - нанкин; 14 - рангу, голова буйвола; 15 - шукин (вариант скрещивания осака и оранда); 16 - тетучио, стальная рыба; 17 - вуалехвост стального цвета; 18 - кинранши (отражаясь от лишенной плавника спины, свет, как бриллиант, горит золотым пламенем); 19 - калико пестрая (гибрид вуалехвоста с пестрым телескопом); 20 - шубункин пестрый с красными пятнами; 21 - голландская пестрая (гибрид голландской львиной головы-оранды с пестрым телескопом); 22 - ханафуса, букет на носу; 23 - осака, голова буйвола; 24 - ватоная, выведенная путем скрещивания вакинХриукин
Иллюстрации, приведенные на с. 202—209, показывают многообразие выведенных селекционерами форм золотых рыбок, гуппи и некоторых других рыб. Выведение золотых рыбок осуществлялось в течение тысячелетий, а селекция гуппи — менее 80 лет (завезена в Европу в начале текущего века). Очень многое сделано селекционерами-аквариумистами в течение последних послевоенных десятилетий. Всего аквариумистами выведено не менее 200 разновидностей рыб разных видов.
Ознакомимся с терминологией
Мы уже не раз употребили термины селекция, порода, предполагая, что большинству читателей эти понятия в общих чертах известны. Теперь рассмотрим подробнее, что понимается под этими терминами, насколько, например, обоснованно многие разновидности декоративных животных, и в частности рыб, считаются породами и не правильнее ли большинство разновидностей декоративных животных и рыб, разводимых любителями-аквариумистами называть не породами, а породными группами, разновидностями, или вариететами.
Селекция — наука о методах сохранения и улучшения существующих и создания новых пород животных и сортов растений, а также гибридов с нужными человеку признаками. Человек стал заниматься селекцией очень давно — тысячи лет назад. Теоретическая основа селекции — генетика — сформировалась как наука только в начале текущего века. К этому времени уже было выведено множество пород животных и сортов растений. Многие породы существуют сотни, если не тысячи лет. Весьма условно методы, применявшиеся селекционерами до становления генетики как науки, можно назвать зоотехническими. В основном они сводятся к интенсивной отбраковке и подбору для разведения пар, в наибольшей степени отвечающих поставленной цели. С генетической точки зрения эти выработанные практикой методы вполне обоснованы и применяются до сих пор.
Под породой в животноводстве понимается созданная с помощью искусственного отбора целостная устойчивая (консолидированная) группа животных одного вида, общего происхождения, имеющих сходные экстерьерно-конституционные, физиологические и хозяйственно полезные признаки, передающиеся по наследству. Породы включают большое число животных, так как только при этом условии возможны творческие отбор и подбор внутри породы, направленные на дальнейшее ее совершенствование.
Численность производителей декоративных животных, включая и рыб, не столь велика, как сельскохозяйственных животных. Их способность передавать свои наследственные свойства потомкам не подвергается достаточно строгой проверке: в подавляющем большинстве случаев декоративные животные недостаточно надежно передают свои ценные качества потомству (нередки «пробросы», а при скрещивании с животными того же вида, но другой окраски или формы в первом поколении получается не однородное по этим признакам качество и т. п.). Поэтому применительно к декоративным животным, включая и рыб, которые не подвергаются необходимым для присвоения «титула» породы испытаниям их наследственных свойств под общественным контролем, более приемлемы термины не порода, а породная группа (предпорода), вариетет, или разновидность.
В животноводстве под породной группой понимается группа домашних животных общего происхождения, отличающаяся специфическими чертами конституции и продуктивности, приспособленностью к определенным пригодно-экономическим условиям, но не обладающая комплексом признаков породы животных. Породные группы составляют обычно помеси, для которых характерны недостаточная выравненность телосложения, продуктивность, племенная ценность (ненадежно передают свои ценные качества потомству). Углубленной племенной работой породные группы улучшают и создают из них высокопродуктивные породы животных.
Термин породная группа очень громоздок и не совсем понятен: при его упоминании напрашивается мысль, что это не предпорода, а группа пород.
В. С. Кирпичников (1979) говорит о породах золотой рыбки, а во втором издании той же книги (1987), то, что ранее именовал породами, он называет разновидностями (расами), то есть заимствует термины, не используемые в животноводстве, но применяемые в биологической литературе. Так же поступил и немецкий исследователь Р. Пихоцкий (1974), назвав свою книгу «Золотая рыбка и ее вариететы». Это обстоятельство делает необходимым пояснить, что биологи называют расой, разновидностью, вариететом.
Раса (от французского race — род, порода) — экологически, поведенчески, а иногда и физиолого-морфологически обособленная совокупность особей (Н. Ф. Реймерс, 1988).
Разновидность — внутривидовая таксономическая категория. В зоологической номенклатуре разновидности приблизительно соответствует вариетет. Вариетет (от лат. varietas — разнообразие, переменчивость) — термин, применяющийся в зоологии к любым подразделениям внутри вида, связанным с изменчивостью — мутацией, возрастными изменениями окраски, географической изменчивостью (Биологический энциклопедический словарь, 1986). В дальнейшем вслед за В. Кирпичниковым (1987) и Р. Пихоцким (1974) мы будем наряду с используемым животноводами термином породная группа применять синонимы разновидность и вариетет, которые, по нашему мнению, более применимы к декоративным животным, разведением которых занимаются любители, а не профессионалы.
Выведенные разновидности декоративных рыбок порою бывают настолько непохожими друг на друга внешне, что неопытный аквариумист принимает их за рыбок различных видов. Например, любители аквариума часто впадают в заблуждение, сталкиваясь с разновидностями (вариететами) золотых рыбок и считая разными видами черных телескопов и оранд или простых золотых рыбок. Такая же путаница происходит при классификации барбусов и цихлид из озер Малави: и Танганьика, дающих межвидовые гибриды.
С чего начать селекцию рыб
Порода, породная группа — это результаты человеческого труда. Выведением пород животных человек начал заниматься в то давнее время, когда приручал первых животных — тысячи лет назад. Если говорить о рыбах, то можно считать, что золотая рыбка была одомашнена во времена Конфуция (551—479 гг. до н.э.), а к периоду династии Мин (1368— 1644)—за несколько сот лет до открытия Г.Менделем законов генетики — были созданы почти все прототипы современных разновидностей золотой рыбки. Таким образом, наши предки не без успеха занимались выведением новых разновидностей животных в течение сотен и тысяч лет, то есть задолго до рождения генетики.
Подавляющее большинство разновидностей аквариумных рыб выведено и продолжает выводиться аквариумистами, мало или совсем не знакомыми с основами генетики. Но из этого не следует, что для аквариумиста-селекционера не нужно знание основных законов общей генетики. Наоборот, использование их может значительно облегчить и ускорить работу по выведению новых разновидностей. Поэтому аквариумистам, желающим серьезно заняться селекцией рыб, необходимо следить за литературой по общей генетике, генетике популяций и частной генетике рыб.
Изучение генетики только по книгам не увлекает и приносит мало пользы. Для того чтобы понять ее законы, научиться использовать их в своей работе, нужно провести ряд скрещиваний и на собственном опыте убедиться в справедливости как минимум трех законов Г. Менделя. Но на это нужно время, а вывести новую породу хочется поскорее. Можно посоветовать аквариумистам начать свою работу, применяя первое время методы, которые условно можно назвать зоотехническими или биотехническими, выделив одновременно несколько (2—3) хотя бы небольших аквариумов для опытов по генетике.
Рассмотрим зоотехнические методы селекции, которыми люди руководствовались до становления генетики как науки, а затем — генетические ее основы. Такой порядок изложения целесообразен и потому, что частная генетика большинства аквариумных рыб изучена пока еще недостаточно, и аквариумисту полезно научиться самому выявлять особенности наследования интересующих его признаков рыбами, которых он разводит.
Начинать попытки выведения новых разновидностей рыб до того, как будет освоено разведение вида, интересующего аквариумиста, не целесообразно. Лишь освоив разведение, ознакомившись с имеющимися в магазинах, у знакомых любителей и на рынке вариететами данного вида, можно поставить себе цель — эталон. Мысленно нужно представлять окраску, форму тела и плавников рыб новой породной группы. Неплохо даже такую, еще не существующую рыбу нарисовать.
Поставленная цель должна быть реальной: необходима гарантия, что в своем хозяйстве или у других любителей удастся подобрать производителей с нужными признаками. Бессмысленно пытаться вывести новую, например, вуалевую породу, если у любителя нет ни одной рыбки с вуалевым хвостовым плавником. Иногда на поиски исходного материала селекционер расходует не один месяц, регулярно посещая зоомагазины, птичий рынок и других аквариумистов.
Чаще всего селекционеру приходится ставить перед собой две задачи:
1. Сохранение и улучшение имеющихся породных групп. Закрепление признаков, обнаруженных им у отдельных или части особей одной и той же разновидности.
2. Выведение новых форм, для чего нужно совмещать признаки двух или более разновидностей: например в одной разновидности селекционеру понравился размер тела рыбы, в другой — окраска, в третьей — форма плавников.
В подавляющем большинстве случаев проще для решения первая задача, поэтому начинать опыты лучше именно с нее. Успех обеспечат настойчивость, целеустремленность, наблюдательность и терпение. Нужно помнить, что для выведения новой разновидности необходимо вырастить несколько поколений, на это уходят месяцы, а иногда и годы упорной работы в одном направлении. Не следует метаться и при неудаче сразу менять направления и цели селекции. Если менять цели — на успех рассчитывать не приходится. При отсутствии перечисленных выше качеств за селекцию лучше не браться.
Совершенствование существующих разновидностей
Если начинающий селекционер научился разводить интересующий его вид рыб, он может заняться улучшением одной из породных групп. Получив потомство от имеющихся производителей, удается выявить признаки, которые желательно закрепить и усилить, и дефекты, от которых желательно избавиться.
Для того чтобы изучить наследственные особенности имеющейся разновидности, необходимо получить не менее двух поколений. Следующей задачей становится отбраковка не отвечающих требованиям (сильно отличающихся от эталона), особей и формирование (подбор) пар производителей, в наибольшей степени соответствующих требованиям эталона. Чем больше производителей отвечают требованиям эталона, тем легче достигнуть успеха в селекции.
Не следует пытаться использовать при разведении как слишком мелких, так и крупных, грубых (по форме тела) и неуклюжих рыб: чаще всего такие попытки обречены на неудачу. Надо ориентироваться на средних.
Важно стремиться к реальной цели — закреплению признаков, которые были замечены неоднократно. Сначала лучше усилить доминантные (господствующие) признаки, а рецессивные (подавляемые) — целеустремленно культивировать в следующих поколениях (доминантные признаки проявляются при скрещивании в первом поколении, а рецессивные — во втором). Лучше выбирать не тех производителей, у которых особенно сильно проявляются отдельные, предусмотренные эталоном признаки, а тех, у которых больше признаков, отвечающих поставленной цели, даже если не все они получили полное развитие.
В подавляющем большинстве аквариумных хозяйств количество производителей (самок и самцов), участвующих в размножении (размножающаяся часть популяции), ограничивается несколькими парами или, в лучшем случае, десятками пар, принадлежащих к разновидности. В среднем, например, самка гуппи за один помет приносит 30 мальков (15 самцов и 15 самок). Для того чтобы ежемесячно получать и выращивать порядка 500 экз. (самцов и самок) гуппи, нужно иметь не более 15—20 пар производителей. При такой маленькой численности размножающейся части неизбежно близкородственное разведение (спаривание родителей с потомками, братьев с сестрами) — инбридинг, приводящий к инбредной депрессии (вырождению), которая выражается в снижении жизнеспособности, темпов роста и окончательных размеров потомства, учащении уродств, уменьшении плодовитости, бесплодии, появлении других нежелательных последствий. У большинства икромечущих для получения 0, 5— 1 тыс. экз. потомков достаточно 2—5 пар хороших производителей.
Чем меньше численность размножающейся части популяции, тем труднее ограничить пагубное влияние близкородственного разведения, тем сильнее будет проявляться инбред-ная депрессия. В то же время, проводя работу по селекции, с неизбежностью приходится умышленно прибегать к наиболее близкородственным спариваниям типа братХсестра, отецХдочь и т. д. Подбор для дальнейшего разведения пар, несущих сходные признаки, также нередко приводит селекционера к бессознательному близкородственному разведению. Этого можно в какой-то степени избежать, если при разведении рыб придерживаться определенной системы, знать родословные производителей. Об этом поговорим несколько позже.
В противоположность инбридингу гетерозис, или гибридная мощность, приводит к превосходству гибридов по ряду признаков и свойств над родительскими формами, которое выряжается в улучшении состояния животных, повышении темпов роста и конечных размеров, жизнеспособности, плодовитости и т. д. Как правило, гетерозис характерен для гибридов только первого поколения, полученных при скрещивании неродственных форм: различных вариететов, линий и даже видов. В дальнейших поколениях (скрещивание гибридов между собой) его эффект ослабляется и исчезает.
Таким образом, чтобы исключить или хотя бы ограничить степень проявления инбредной депрессии, нужно стремиться к тому, чтобы избежать в доступных пределах близкородственного разведения. Но как это сделать? Ведь проблема борьбы с инбредной депрессией — одна из наиболее сложных проблем не только аквариумного, но и промышленного рыбоводства.
Аквариумисту можно рекомендовать два способа ограничения инбредной депрессии:
1. Увеличение численности размножающейся части популяции (пар производителей) до необходимых пределов, содержание всех их вместе для строго случайного спаривания. При этом количество самцов и самок должно быть одинаковым. По мнению большинства исследователей, количество участвующих в размножении особей обоего пола должно исчисляться сотнями экземпляров. Р. Р. Стикни (1986), ссылаясь на Х.Л.Кинкайда (1976, 1977) и указывая меньшую численность производителей (50—100, но не менее 25 пар), оговаривается, что и в этом случае инбридинг полностью не исключается. Для борьбы с этим явлением возникает необходимость в периодическом приобретении новых производителей. Использование этого метода ограничивает возможность строгой отбраковки особей, не отвечающих требованиям селекционера.
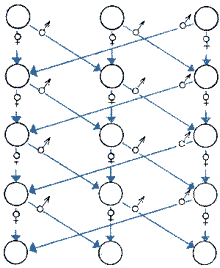
2. Разведение по линиям. Линия — группа родственных особей, характеризующихся определенными признаками, постоянно воспроизводящимися в ряду поколений. При этом подразумевается генотипическая однородность линии по генам, контролирующим эти признаки.
Чтобы получить сильное потомство, нужно содержать и систематически использовать при разведении животных не менее двух родственных линий производителей. Но наиболее приемлемым, по мнению Р. Р. Стикни (1976), является чередующееся скрещивание трех линий. Степень инбридинга при этом несколько меньше, чем при случайном спаривании. Такая схема решает проблему замены производителей, возникающую при использовании только двух размножающихся линий. Минимальное количество пар производителей в этом случае не оговаривается. По всей вероятности, оно может быть небольшим.
Чередующиеся линии получают разделением животных, которые дают потомство, на три группы А, В и С. Во время каждого периода спаривания самцы линии А скрещиваются с самками линии В, самцы линии В — с самками линии С, самцы линии С — с самками линии А.
Для гарантии запланированного скрещивания необходимо каждую линию содержать отдельно. Для живородящих рыб, кроме этого, обязательно раздельное содержание до спаривания самцов и самок.
| ЛинияA | Линия B Генерация |
ЛинияC | |
0
1
2
3
4 |
 |
||
| Схема чередующегося трехлинейного скрещивания для уменьшения инбридинга в популяциях рыб | |||
При любой системе разведения одним из самых ответственных моментов в работе селекционера, независимо от того, стремится он сохранить или улучшить имеющуюся у него породную группу или вывести новую, является отбраковка самцов и самок, не отвечающих требованиям эталона (поставленной цели), и подбор лучших производителей для дальнейшего разведения (на племя). В большинстве случаев целесообразно отбраковывать в течение всего периода выращивания самцов и самок. В этом случае подбор пар [Производителей производится в конце периода выращивания из оставшихся рыб, не имеющих бросающихся в глаза дефектов.
При селекции видов рыб, у которых ярко выражены сцепленные с полом признаки (например, гуппи), выбор самок существенно облегчается после содержания их в растворе мужских половых гормонов (например, в растворе метилтестостерона), под влиянием которых проявляются некоторые, не заметные в обычных условиях, сцепленные с женским полом признаки окраски тела и плавников.
Гормональная обработка самок некоторых рыб облегчает работу селекционера, сокращая объем работ по проверке производителей по потомству. Но так как далеко не всегда лучшие по фенотипу (внешним признакам) производители способны давать такое же хорошее потомство, наиболее аффективным методом подбора производителей является оценка их по качеству потомства. Для этого спермой одного самца осеменяют икру нескольких самок; потомков, полученных от разных самок, выращивают отдельно. Этот метод наиболее эффективен, но очень трудоемок и требует использования большого количества аквариумов.
Вообще, имея всего 2—3 аквариума, редко можно рассчитывать на успех в проведении селекционной работы даже в наиболее простом ее варианте, ограничивающемся совершенствованием только лишь одной породной группы.
Выведение новых разновидностей
Когда исходный материал подобран, приступают к скрещиванию самок и самцов из разных породных групп. Здесь следует повторить, что результаты, полученные только в одном первом поколении, в большинстве случаев ни о чем не говорят, так как в первом поколении проявляются только доминирующие признаки. Начиная с первого поколения, закрепляются и становятся заметными только признаки, сцепленные с мужским полом, они четко переходят от отца к сыну из поколения в поколение. Для закрепления не сцепленных с полом или сцепленных с женским полом признаков предстоит еще большая работа и для ее выполнения потребуется вырастить несколько поколений.
| Воспроиз- водительное |
Вводное | Поглоти- тельное |
Альтерна- тивное |
|
|---|---|---|---|---|
| P | AxB | AxB | AxB | AxB |
| F1 | F1 | AxF1 | F1xB | AxF1 |
| F2 | F2 | AxF2B | F3BxB | F2BxB |
| F3 | F3 | AxF3B | F3xB | AxF3B |
| P - родительское поколение; F1, F2,F33 - соответственно первое, второе, третье поколения |
||||
О массовом и индивидуальном отборе, а также об инбридинге на выдающегося производителя уже говорилось. Теперь остановимся на других специальных типах скрещивания.
При выведении новых разновидностей применяют разные типы скрещивания. Основные из них: вопроизводительное, вводное, поглотительное и альтернативное.
1. Воспроизводительное скрещивание применяется, когда надо сочетать многие полезные признаки обеих скрещиваемых разновидностей. Такое скрещивание требует очень тщательной селекции во всех гибридных поколениях (под селекцией в данном случае понимается отбраковка и подбор производителей для дальнейшего разведения).
2. Вводное скрещивание используют, когда в имеющуюся разновидность хотят ввести один или немногие признаки другого вариетета. Каждое поколение гибридов скрещивают возвратно с особями имеющейся (собственной) разновидности. При этом необходимо сохранять на племя возвратных гибридов, имеющих исходный признак породы-улучшателя. Если этот признак определяется одним или немногими доминантными генами с четким проявлением, проблема сохранения нужного признака решается сравнительно легко. При рецессивности генов, и тем более при полигенном (определяемом многими генами) наследовании, опасность утери признака-улучшателя очень велика.
3. Поглотительное скрещивание по характеру похоже на вводное. После исходного скрещивания двух разновидностей ставят серию возвратных скрещиваний, но гибриды повторно скрещивают с особями породы-улучшателя, а не имеющейся (собственной) разновидности.
Вводное (поглотительное) и воспроизводительное скрещивание часто сочетают друг с другом.
4. Альтернативное (чередующееся) скрещивание, по мнению В. С. Кирпичникова (1979, 1987), позволяет в наибольшей степени избежать инбридинга при сочетании признаков двух разновидностей. Попеременное скрещивание гибридов с особями двух вариететов сопровождают отбором нужных комбинаций признаков. Через 3—4 поколения альтернативное скрещивание заменяют воспроизводительным, так как в противном случае трудно добиться стабилизации признаков новой гибридной разновидности.
Не следует путать пользовательное (промышленное) скрещивание — получение интересных гибридов первого поколения с выведением не только новых пород, но даже и разновидностей. Такие гибриды могут быть очень привлекательными, стать украшением аквариума, но их нельзя отождествлять с результатами завершенной работы по выведению новой породы или разновидности. По существу в большинстве случаев это использование гетерозиса, проявляющегося только в одно первом поколении.
Выведение новых форм путем скрещивания разных пород одного и того же вида, о котором шла речь выше, называется внутривидовым в отличие от межвидового и межродового скрещиваний.
В отличие от млекопитающих не только межвидовые, но и межродовые гибриды рыб иногда оказываются плодовитыми, поэтому в аквариумном рыбоводстве гибридизация является одним из перспективных методов выведения новых разновидностей. Достаточно сказать, что все цветные меченосцы были получены путем скрещивания меченосцев и пецилий (X. helleri x X. maculatus). К сожалению, аквариумисты этому методу выведения новых форм декоративных рыб уделяют явно недостаточное внимание. Хочется надеяться, что гибридизация и пользовательное скрещивание в перспективе займут достойное место в практике аквариумного рыбоводства. Ведь для любителей декоративного аквариума безразлично— чистопородные или помеси, плодовитые или бесплодные рыбы украшают их аквариум.
В заключение еще один, последний, совет. Для успешной работы по селекции необходимо вести дневник наблюдений, в котором записывать все интересные наблюдения, случаи и изменения, происходящие в аквариумном хозяйстве. Кроме этого, нужно вести дневник пересадок и в нем регистрировать, когда, сколько и откуда посажены рыбы в тот или иной аквариум с указанием их происхождения. Вдобавок к дневнику пересадок на каждый аквариум хорошо повесить табличку. В ней отмечают: дату посадки, какая рыба посажена, даты нереста, сколько мальков родилось и т. д. Это оперативная информация. Таблички можно переносить вместе с рыбой с одного аквариума на другой. Нужно также, по возможности подробнее, вести упрощенные родословные производителе (самок и самцов). В них делают записи о происхождении рыбы с краткой характеристикой основных признаков (форм плавника, основная и покрывающая окраска и т.п.), с кем скрещивалась (самка или самец), сколько получено потомство даты получения пометов, характеристика потомков и т. д.
При выведении новой разновидности желательно вести специальный журнал, в который записывать план выведения вариетета, характеристику исходных породных групп, результаты скрещиваний, «знаменательные» даты.
Конечно, на такие записи уходит много времени, но впоследствии это сполна окупается. Такой «багаж» поможет составить собственное представление о характере наследования отдельных признаков, а это значительно облегчит в дальнейшем работу по выведению новых разновидностей, избавит от ошибок, на исправление которых уходит иногда, намного больше времени, чем на ведение записей.
Генетические основы селекции
Генетика — наука о наследственности и изменчивости живых организмов и методах управления ими. В ее основу легли закономерности наследственности, обнаруженные Г. Менделем при скрещивании различных сортов гороха (1865), а также мутационная теория X. Де Фриза.
Как уже говорилось, аквариумисту, решившему заняться селекцией рыб, желательно знать основные законы общей генетики и ознакомиться с частной генетикой рыб. Законы общей генетики справедливы для всех видов растений и животных, включая человека, а частная генетика изучает особенности передачи по наследству разных признаков конкретного вида. Можно говорить о частной генетике золотой рыбки, макропода, гуппи и т. д. Так как частная генетика подавляющего большинства обитателей наших аквариумов изучена очень слабо, аквариумисту-селекционеру приходится собственными силами изучать особенности наследования интересующих его признаков своими питомцами. Для этого как минимум необходимо знать основы общей генетики и уметь использовать простейшие методы гибридологического анализа.
Прежде всего восстановим в памяти терминологию.
Ген — наследственный фактор, материальная единица наследственности, ответственная за формирование какого-либо признака. Входит в состав хромосом. Контролируя образование белков (ферментов и др.), гены управляют всеми химическими реакциями организма и таким образом определяют его признаки.
Признак — какое-то отдельное качество организма, по которому можно отличать один организм от другого.
Наследственные признаки — признаки, в норме присущие особям данного вида или вновь возникшие в результате изменения генотипа (мутации) и воспроизводящиеся при размножении во всех последующих поколениях.
Мутации — наследуемые изменения генетического материала, внезапные, естественные или вызванные искусственно, приводящие к изменению тех или иных признаков организма. Именно мутации дают основной материал для естественного и искусственного отбора, являясь необходимым условием эволюции в природе и выведения новых разновидностей животных и растений.
Приобретенные признаки (модификации) — признаки, отсутствующие у предков данной особи и приобретенные организмом в течение его индивидуальной жизни. Приобретенные признаки не связаны с изменением генотипа, поэтому не наследуются потомством. Они возникают под влиянием специфических условий среды. Пределы этих изменений контролируются генотипом.
Признаки качественные (альтернативные, олигогенные) — признаки, контролируемые одним или немногими генами, действие которых отчетливо отграничено от действия ненаследственных факторов. Различия по качественным признакам устанавливаются непосредственно путем наблюдения или сравнения, без измерения или взвешивания. Они обладают прерывной изменчивостью и описываются по принципу «есть-нет»; например, красный, синий корпус у самца гуппи — есть у него пятно на груди или нет и т. д. Степень проявления качественных признаков почти не зависит от влияния среды.
Признаки количественные (мерные, полигенные) — признаки, генетически контролируемые суммарным действием большого числа генов (полигены). Количественные признаки могут приобретать цифровое выражение, которое может устанавливаться не только глазомерно (большой, маленький), но и путем взвешивания, подсчета. Вследствие полигенного контроля и большой модификационной изменчивости под влиянием внешней среды количественные признаки обладают непрерывной изменчивостью. Они могут быть описаны по принципу «больше-меньше», то есть один и тот же признак, присущий разным особям или формам, имеет различную степень выражения. К количественным признакам относятся плодовитость рыб, число лучей в плавниках и др. Проявление количественных признаков в значительной степени зависит от условий внешней среды (температуры, характера питания и т. п.).
По характеру наследования признаки можно разделить на доминантные и рецессивные.
Признак доминантный — признак из пары противоположных признаков (например, окраска рыб, свойственная неодомашненным особям), проявляющийся у гетерозиготных особей вследствие подавления доминантным аллелем (А), определяющим развитие доминантного признака, рецессивного аллеля (а), ответственного за проявление противоположного — рецессивного признака. Например, дикая серая окраска гуппи доминирует над светлой.
Признак рецессивный — признак, не проявляющийся у гетерозиготной особи вследствие подавления действия рецессивного аллеля (а), контролирующего развитие рецессивного признака действием доминантного аллеля (А) этой же аллельной пары. Рецессивный признак проявляется только в том случае, если контролирующий его рецессивный аллель находится в гомозиготном состоянии. Например, альбинизм (красные глаза, отсутствие темного пигмента) рецессивен у всех животных, включая и рыб.
Кодоминантность — участие обоих аллелей в определении признака у гетерозиготной особи; частный случай доминантности.
Генотип — совокупность аллелей гена или группы генов, контролирующих анализируемый признак у данного организма (в этом случае нерассматриваемая часть генотипа выступает в качестве генотипической среды). Генотип контролирует развитие, строение и жизнедеятельность организма, то есть совокупность всех признаков организма — его фенотип. Особи с разными генотипами могут иметь одинаковый фенотип, поэтому для определения генотипа организма необходимо проводить его генетический анализ, например анализирующее скрещивание. Особи с одинаковым генотипом в различных условиях могут отличаться одна от другой по характеру проявления признаков (особенно количественных), то есть различаться по фенотипу. Таким образом, генотип определяет возможные пути развития организма и его отдельных признаков во взаимодействии с внешней средой (например, гирардинус, выращенный при низких температурах, оказывается темнее своих братьев и сестер, живущих при более высоких температурах).
Фенотип — совокупность всех признаков и свойств особи, формирующихся в процессе взаимодействия генотипической структуры (генотипа) и внешней по отношению к ней среды. В фенотипе не реализуются все генотипические возможности, и он является лишь частным случаем реализации генотипа в конкретных условиях. Поэтому даже между однояйцевыми близнецами, имеющими полностью идентичные генотипы, можно выявить заметные фенотипические различия, если они развивались в разных условиях.
Аллели (аллеломорфы, аллельные гены) — формы состояния одного и того же гена, находящиеся в гомологических участках (локусах) гомологичных хромосом и контролирующие развитие альтернативных признаков. Два аллеля у диплоидных организмов не могут находиться в одной гамете. Аллель — одно из возможных структурных состояний гена — определяет вариант развития одного и того же признака организма. Возникает при любом изменении структуры гена в результате мутаций или за счет внутригенных рекомбинаций (возможное число аллелей каждого гена неисчислимо). Наличием аллельных генов обусловлены фенотипические различия среди особей.
Гаметы — зрелые мужские сперматозоиды и женские (яйцеклетки) половые клетки, содержащие гаплоидное (половинное) вследствие редукции в мейозе число хромосом.
Зиготы — оплодотворенные яйцеклетки. Если сливаются обычные, редуцированные гаметы, зигота имеет двойное (диплоидное) число хромосом.
Гомозигота — диплоидная клетка или особь, гомологичные хромосомы которой несут идентичные аллели того или иного гена.
Гетерозигота — клетка или особь, у которой гомологичные хромосомы несут различные аллели (альтернативные формы) того или иного гена.
Ознакомление с перечисленными выше терминами позволит в более сжатом виде изложить те законы генетики, которые полезно вспомнить аквариумисту, решившему заняться селекцией рыб. Теперь перейдем К рассмотрению основных законов генетики.
| P- родители | 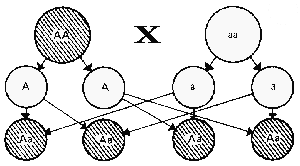 |
| Гаметы | |
| F1 - первое поколение | |
| Схема, иллюстрирующая единообразие гибридов первого поколения: А - доминантный ген; а - рецессивный ген; заштрихованный круг - доминантный фенотип (дикая окраска); светлый круг - рецессивный фенотип (альбиносы) | |
Закон единообразия гибридов первого поколения (первый закон Г. Менделя) утверждает, что потомство первого поколения от скрещиваний устойчивых форм, различающихся по одному признаку, имеет одинаковый фенотип по этому признаку. При этом все гибриды могут иметь фенотип одного из родителей (полное доминирование) или промежуточный фенотип (неполное доминирование), кроме этого, гибриды могут проявить признаки обоих родителей (кодоминирование). Этот закон основан на том, что при скрещивании двух гомозиготных по разным аллелям форм (АА и аа) все их потомки одинаковы по генотипу (гетерозиготы — Аа), а следовательно, и по фенотипу.
Иллюстрировать этот закон можно следующим примером: если скрестить рыбу любого вида, имеющую дикую окраску, с рыбой того же вида — альбиносом (основной признак альбиноса — красные глаза, даже белая особь не с красными глазами не может считаться альбиносом), то в первом поколении мы получим всех без исключения потомков имеющих доминантную дикую окраску. Если в первом поколении часть потомков — альбиносы, то родитель, имевший дикую окраску, был гетерозиготен.
Альбинизм у всех видов животных рецессивен. И при скрещивании с доминантами он, как все рецессивы, может появиться только во втором поколении.
Скрещивание обозначают в генетике знаком умножения (X). При написании схемы скрещивания принято на первом месте ставить женский пол. Женский пол обозначают знаком 9 (зеркало Венеры), мужской— (щит и копье Марса). Родительские организмы, взятые в скрещивание, обозначают буквой Р. Потомство от скрещивания других особей с различной наследственностью называют гибридным, а отдельную особь — гибридом. Гибридное поколение обозначают буквой F с цифровым индексом, соответствующим порядковому номеру гибридного поколения (F1, F2, F3 и т.д.). Гибридов, получаемых от скрещивания особей, различающихся по некоторым признакам, но относящихся к одному виду, называют внутривидовыми (иногда метисами, или помесями). Отдаленных гибридов, происшедших от скрещивания особей из разных видов и родов — соответственно межвидовыми и межродовыми.
| Гаметы F1 | 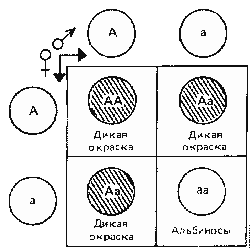 |
| F2 | |
| Схема, иллюстрирующая закон расщепления признаков у потомства второго поколения (F2). Альбиносы (аа) и гомозиготные особи с дикой серой окраской (АА) составляют по 25% общего количества гибридов, а 50% - серые гетерозиготы (Аа) | |
Закон расщепления (второй закон Г. Менделя) гласит, что при скрещивании гибридов первого поколения между собой среди гибридов второго поколения в определенных соотношениях появляются особи с фенотипами исходных родительских форм и гибридов первого поколения. В случае полного доминирования выявляются 75% с доминантным и 25% с рецессивным признаком, то есть два фенотипа в соотношении 3 : 1. При небольшом количестве особей во втором поколении по теории вероятности от указанных соотношений могут наблюдаться существенные отклонения. Например, если потомков всего 4, то в их числе может не оказаться ни одного альбиноса (аа).
Существенные отклонения от указанных соотношений могут наблюдаться, если жизнеспособность гамет и зигот всех типов неодинакова. Так, например, альбиносы менее жизнеспособны по сравнению с особями, имеющими дикий генотип; их может оказаться значительно меньше, чем предполагалось по расчету.
При неполном доминировании и кодоминировании 50% гибридов второго поколения (F2) имеют фенотип гибридов первого поколения и по 25% — фенотипы исходных родительских форм, то есть наблюдается расщепление 1 : 2 : 1.
В основе второго закона лежит закономерное поведение пары гомологичных хромосом (с аллелями А и а), которые обеспечивают образование у гибридов первого поколения гамет двух типов, в результате чего среди гибридов второго поколения выявляются особи трех возможных генотипов в соотношении 1АА: 2Аа: 1аа.
Конкретные типы взаимодействия аллелей и дают расщепление по фенотипу в соответствии со вторым законом Г. Менделя.
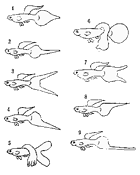
Для того чтобы лучше понять суть первого и второго законов Г. Менделя, приведем рассказ известного генетика, аквариумиста Федора Михайловича Полканова (1970). Как-то он обнаружил в рыборазводне Московского зоопарка среди обычных барбусов суматранусов трех альбиносов с золотым телом, красными глазами и еле просвечивающимися темными полосами. Основные признаки альбиносов—красные глаза и неспособность вырабатывать темные пигменты. Пропадала полностью или почти полностью (неполный альбинизм) черная окраска, и рыба стала светлой без черных полос и пятен, желтой, золотистой, светло-зеленой, белой. Федор Михайлович взял альбиносов себе и вырастил. Все оказались самцами. Одного из самцов он скрестил с обычной, не золотой, самкой. Теоретически можно было ожидать три разных типа наследования. Все потомки от измененной рыбы (в данном случае от альбиноса) по виду могли оказаться измененными. Это означало бы, что измененный признак является доминантным. Второй случай — все потомки промежуточные. И третий — когда измененный признак не проявился ни у одного из потомков, иначе говоря, оказался рецессивным. Могло оказаться, что в потомстве обнаружатся и нормальные суматранусы с дикой серой окраской и альбиносы. Но это в том случае, если обычная, не золотая, самка оказалась бы гетерозиготной, то есть помесью обычного суматрануса с альбиносом.
Альбиносы обнаружены почти у всех видов рыб, встречаются они и у пресмыкающихся, и у птиц, и у млекопитающих, и даже у человека. Часто альбиносы менее жизнестойки и менее плодовиты, хуже растут. Но не всегда. Альбинизм - признак рецессивный.
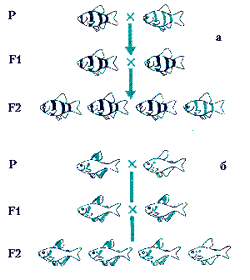
Если родительские формы наследственно чисты, то в соответствии с первым законом Г. Менделя в первом поколении все потомки одинаковы, поэтому этот закон и называют законом единообразия первого поколения.
В опыте Федора Михайловича в первом поколении все барбусы, как и следовало ожидать, оказались обычными — ни одного золотого. Те, кто забыл первый закон Г. Менделя, разочаровались бы и прекратили опыт. Но Ф. М. Полканов был не только опытным аквариумистом, но и генетиком. Он понимал, что заинтересовавший его признак не пропал. Он вырастил потомков золотого самца и скрестил между собой. Часть мальков (примерно 1/4) оказалась альбиносами, а 3/4 — обычными барбусами. Три к одному. Это соотношение полностью соответствует второму закону Г. Менделя — закону расщепления, разделения гибридов второго поколения на исходные формы.
Еще несколько примеров. Все мы знаем барбусов семифасциолятусов и барбусов шуберти и думаем, что это разные виды. А оказывается это один и тот же вид. Ярко-лимонные шуберти возникли в результате мутации одного гена у барбуса семифасциолятуса. Немецкому аквариумисту удалось закрепить этот признак, и лимонные семифасциолятусы, мало похожие на своих предков, были описаны как отдельный вид и названы по фамилии выведшего их аквариумиста. Барбус семифасциолятус и барбус шуберти — очень хорошие объекты для того, чтобы убедиться в справедливости первого и второго законов Г. Менделя. Их нетрудно развести. Скрестите их. Первое поколение не будет отличаться от семифасциолятусов, а во втором — примерно 75% окажутся обычными семифасциолятусами, а 25% — шуберти.
Мало кто из аквариумистов не знает хифессобриконов серпас и минор. Оказывается это тоже один и тот же вид, минор — рецессивная форма, рецессивный вариант того же гена, который определяет окраску серпаса. Серпас обитает в реках Бразилии — в мутновато-коричневой воде. Мутация, в результате которой возник минор, позволила хифессобрикону расширить свой ареал и заселить участки реки с более светлой водой, на красноватой глинистой почве.
Если скрестить серпаса и минора, то как и в случае с барбусом, в первом поколении все потомки будут похожи на серпаса, а во втором — будет получено примерно 25 % миноров и 75% серпасов.
Если скрестить серых гуппи со светлыми, то доминирующей будет серая фоновая окраска, рецессивной — светлая. В первом поколении будут получены только серые особи, во втором — примерно 75% серых и 25% светлых.
Вернемся к суматранусам. Для выведения альбиносов правильнее было бы скрещивать не рыб первого поколения между собой, а самок первого поколения с исходными золотыми самцами — их отцами (инбридинг на выдающегося производителя). В этом случае рецессивных потомков получилось бы не 25%, а 50%. Такой тип скрещивания называется анализирующим, но он полезен не только для анализа, но и с селекционными целями.
 Результаты скрещивания: а - обычного барбуса суматрануса с альбиносом; б - хифессобриконов серпаса с минором
Результаты скрещивания: а - обычного барбуса суматрануса с альбиносом; б - хифессобриконов серпаса с минором
Закон независимого комбинирования (наследования) признаков (третий закон Г. Менделя) утверждает, что каждая пара альтернативных признаков ведет себя в ряду поколений независимо одна от другой, в результате чего среди потомков второго поколения в определенном соотношении появляются особи с новыми (по отношению к родительским) комбинациями признаков. Например, при скрещивании исходных форм, различающихся по двум признакам, во втором поколении выявляются особи с четырьмя фенотипами в соотношении 3 : 3 : 1 (случай полного доминирования). При этом два фенотипа имеют родительские сочетания признаков, а остальные два — новые. Этот закон основан на независимом поведении (расщеплении) нескольких пар гомологичных хромосом. Так, при дигибридном скрещивании это приводит к образованию у гибридов первого поколения четырех типов гамет (АВ, Ав, аВ, ав) и после образования зигот — к закономерному расщеплению по генотипу и соответственно по фенотипу.
Для селекционера важно знать, что чем больше он хочет совместить признаков, взятых из разных вариететов, тем труднее решить эту задачу, так как с увеличением количества совмещаемых признаков возрастает количество потомков второго поколения, которое необходимо выращивать, и выбрать для дальнейшей работы необходимое количество пар особей с необходимыми генотипами и фенотипами (табл. 10).
 |
|
| Родительский генотип P | 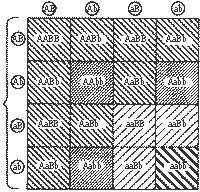 |
| Гаметы | |
| Первое поколение F1 | |
| Второе поколение F2 | |
| Схема, иллюстрирующая независимое комбинирование признаков. Наследование дикой (серой - B)и светлой (b) основной окраски тела у гуппи, а также нормальных размеров (A) и гигантизма (a). Генотипы родителей и потомков обозначены комбинацией букв, а фенотипы - штриховка. | |
Допустим, что поставлена задача вывести гуппи-гигантов со светлой основной (фоновой) окраской.
В нашем примере при дигибридном скрещивании мы имели четыре типа гамет АВ, Ав, аВ и ав; 16 их возможных комбинаций (42=16), число фенотипических классов— четыре (серые нормального размера, серые гиганты, светлые нормального размера и светлые гиганты) при девяти генотипических классах, из которых на долю гигантов пришелся только один. Если мы хотим вывести породную группу светлых гигантов, то, для того чтобы подобрать одну пару производителей, нужно иметь более 32 гибридов второго поколения (в том числе 16 самок и 16 самцов).
Установленные законом Г. Менделя в их классической форме соотношения проявляются не всегда.
Для выявления законов Г. Менделя в их классической форме необходимы:
- гомозиготность исходных форм;
- образование у гибридов гамет всех возможных типов в равных соотношениях, что обеспечивается правильным течением мейоза (редукционного деления клеточного ядра, предшествующего образованию половых клеток);
- одинаковая жизнеспособность гамет всех типов;
- равная вероятность встречи любых типов гамет при оплодотворении;
- одинаковая жизнеспособность зигот всех типов.
| 1 -AABB +2 - AABb +2 - AaBB +4 - AaBb 9 |
Серые нормального размера | 1 -AAbb +2 - Aabb 3 |
Светлые нормального размера | 2 - aaBb +1 - aaBB 3 |
Серые гиганты |
| 1 -aabb
1 |
Светлые гиганты |
| Число пар аллелей | Гаметы | Число классов | ||
|---|---|---|---|---|
| число типов | число комбинаций | фенотипических при полном доминировании | генотипических | |
| 1 | 2 | 4 | 2 | 3 |
| 2 | 4 | 16 | 4 | 9 |
| 3 | 8 | 64 | 8 | 27 |
| 4 | 16 | 256 | 16 | 81 |
| 5 | 32 | 1024 | 32 | 243 |
| 6 | 64 | 4096 | 64 | 729 |
| ... | ||||
| n | 2n | 4n | 2n | 3n |
Нарушение этих условий может приводить либо к расщеплению в первом поколении, либо к искажению соотношения различных генотипов и фенотипов.
Кроме этого, нарушение соотношений, вытекающих из третьего закона Г. Менделя, наблюдается в случаях сцепления генов (явление, в основе которого лежит локализация генов в одной хромосоме, оно выражается в том, что аллели сцепленных генов, находящихся в одной группе сцепления, имеют тенденцию наследоваться совместно). Это приводит к образованию у гибрида гамет преимущественно с родительскими сочетаниями аллелей.
В целом законы Г. Менделя справедливы для аутосомных (обычных, не половых) генов с полной пенетрантностью (частотой фенотипического проявления гена в популяции особей, являющихся его носителями). При полной пенетрантности доминантный или рецессивный гомозиготный аллель проявляется у каждой особи, в генотипе которой он имеется, с постоянной экспрессивностью (силой действия гена, характеризующейся степенью фенотипического проявления признака, контролируемого данным геном). Пенетрантность и экспрессивность зависят от взаимодействия данного гена с внешними условиями и генотипической средой (действием других генов).
Вспомнив законы Г. Менделя, можно приступить к изучению особенностей наследования интересущих признаков.
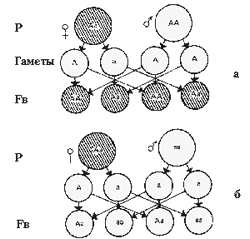 Бекросс: а - возвратное скрещивание гибрида F1 с доминантной родительской формой; б - анализирующее скрещивание (возвратное скрещивание гибрида F1 c рецессивной родительской формой)
Бекросс: а - возвратное скрещивание гибрида F1 с доминантной родительской формой; б - анализирующее скрещивание (возвратное скрещивание гибрида F1 c рецессивной родительской формой)
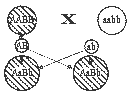
Для анализа характера наследования признаков (выявления характера доминирования и др.) применяется генетический (гибридологический) анализ. Классическая схема его предусматривает выделение исходных гомозиготных форм, получение от них гибридов первого поколения (F1) и скрещивание их (F1) между собой — получение гибридов второго поколения (F2). Однако для генетического анализа более эффективно скрещивание гибрида с одной из родительских форм, несущей данную пару аллелей в гомозиготном состоянии (возвратное — реципрокное скрещивание — беккросс). Беккросс обозначается как FB.
Все гаметы родительской формы будут нести доминантную аллель А, а у гибридов образуются гаметы двух сортов — с аллелями А и а. Поэтому в результате случайного сочетания этих гамет при оплодотворении в потомстве от такого скрещивания имеет место расщепление по генотипу в соотношении 2Аа : 2АА, или 1 : 1, в то время, как расщепления по фенотипу не произойдет (1 : 0) — все особи в FB будут иметь дикую окраску.
Значительно больший интерес для генетического анализа представляет скрещивание гибрида F1 (Аа) с формой, гомозиготной по рецессивному гену (аа). Поскольку все гаметы этой формы несут рецессивную аллель, характер расщепления в потомстве по фенотипу будет соответствовать качеству гамет гибридного организма. В результате расщепление по гену окраски рыб окажется в соотношении 1Аа : 1аа. По характеру расщепления в потомстве от такого скрещивания Аа Х аа можно проанализировать наследственную структуру гибрида по данному гену.
С помощью такого скрещивания из гибридов первого и из любого последующего поколения можно отобрать особей, гомозиготных по доминантному гену, от особей гетерозиготных.
Благодаря этому, скрещивание организма с исходной формой, гомозиготной по рецессивному гену, получило название анализирующего скрещивания. Как уже указывалось (пример с барбусом), такой тип скрещивания полезен и при выведении новых разновидностей.
Выше мы рассмотрели самое простое, главное, что необходимо знать аквариумисту, решившему заняться селекцией рыб. В практике встречаются значительно более сложные случаи и отклонения от законов Г. Менделя, исключения, которые, если с ними получше разобраться, оказываются не опровергающими, а, наоборот, подтверждающими эти фундаментальные законы. Из-за недостатка места мы не смогли обогатить текст достаточным количеством примеров и иллюстраций, рассмотреть особенности наследования при различных типах взаимодействия генов, сцепленное с полом наследование, влияние факторов внешней и внутренней (генотипической) среды на действие генов и т. д. Все это накладывает отпечаток на результаты скрещиваний, приводит к отклонениям от ожидаемых теоретических результатов. Но даже и знание того, что удалось осветить в данной главе, может оказать существенную помощь в понимании результатов скрещиваний по какой бы системе они не проводились.
В заключение не мешает напомнить, что подавляющее большинство интересующих селекционера признаков проявляется в полной мере только при оптимальных условиях содержания и кормления рыб.